
TEORIA EVOLUCIONISTA
OU DA
GERAÇÃO ESPONTÂNEA GRADUAL
Não sendo possível conceber qualquer modelo teórico para a origem da vida fora da realidade que conhecemos, que é a da Terra, é neste contexto que a maioria dos cientistas procuraram encontrar a resposta à questão.
A abordagem do problema não pode, por outro lado, ser desinserida da perspectiva evolucionista que Charles Darwin (1809-1882) nos legou. Sabendo que as espécies se formaram por evolução a partir de outras, menos complexas, a lógica conduz-nos a focar a nossa atenção nas formas de vida mais simples e a colocar a questão fundamental a esse nível: como se formaram os microrganismos procariontes ? Thomas Huxley terá sido um dos primeiros a esboçar a ideia ousada que consistia em ligar de forma contínua o não vivo ao vivo. Faltavam-lhe, todavia, elementos para prosseguir. Estas ideias foram retomadas, já em pleno século XX, por J. B. S. Haldane (1892-1964) e A. I. Oparin.
Estes dois cientistas, sem se comunicarem, formularam a hipótese de que a atmosfera primitiva era, não só diferente pela sua composição, da actual mas, sobretudo, que não seria oxidante; o oxigénio, a existir, teria destruído qualquer formação pré-biótica. Nesta hipótese base, assenta a Teoria evolucionista da origem da vida ou da geração espontânea gradual. Consiste, basicamente, em admitir que, de forma espontânea e gradual, em condições ambientais diferentes das que actualmente existem, entidades moleculares se tenham combinado, dando origem, primeiro, a novas moléculas (que nós classificamos de orgânicas), e depois, a moléculas mais complexas por justaposição das primeiras (os polímeros). Finalmente, estruturas moleculares complexas e constituindo entidades isoladas do meio, com capacidades metabólicas e de reprodução, sujeitas às leis da evolução (diversidade-selecção-evolução), terão estado na origem das primeiras células vivas.
O ambiente primitivo
Sabe-se que a atmosfera da Terra e as condições ambientais que nela reinam hoje são completamente distintas daquelas que imperavam há cerca de 4,6 biliões de anos, quando ocorreu a condensação do sistema solar a partir de uma nuvem de gases e poeiras.
As condições então reinantes, ainda que muito diferentes das actuais, satisfaziam, em princípio, os requisitos para o desenvolvimento de formas vivas. Em primeiro lugar, a Terra localizava-se próximo de uma fonte de energia, o Sol. Em segundo lugar, dos nove principais planetas em órbita solar, era a massa terrestre a que se não encontrava tão perto dele que os seus elementos constituintes lhe fossem arrebatados sob a forma de gases, ou liquefeitos em rocha fundida. Também não era tão grande a distância, que os seus gases congelassem, tal como actualmente sucede em Titã, a maior lua de Saturno. A água é líquida na Terra, mas não em Mercúrio, onde foi toda evaporada para o espaço, ou em Júpiter, onde ocorre sob a forma de gelo. Por fim, a Terra era um corpo suficientemente grande para conservar a atmosfera, possibilitando o ciclo fluido dos elementos, mas não tanto que a sua gravidade mantivesse uma atmosfera demasiado densa, impeditiva da passagem dos raios solares.
A origem da vida na Terra ou a sua inoculação por seres extraterrestres terá ocorrido no período entre 3,9 e 2,5 biliões de anos, conhecido por período Arqueano. Com efeito, os registos fósseis mais recuados que se conhecem datam de há 3,5 biliões de anos.
Qual seria a conformação da Terra e a composição da sua atmosfera, nesse período? Eis a primeira questão à qual importa responder, para se conhecer o cenário em que se terão desenrolado os fenómenos subjacentes ao aparecimento da vida.
A Terra estaria ainda em fase de arrefecimento. A crosta terrestre era necessariamente frágil e os fenómenos de vulcanismo, frequentes. Frequente era também o bombardeamento por diversos corpos celestes, cometas e meteoritos. Com o arrefecimento da Terra, parte da água trazida à superfície pelas erupções vulcânicas e pela desgaseificação de minerais condensou-se e formou os mares primitivos (a hidrosfera). Outra parte, conjuntamente com os gases, foram conferindo à atmosfera uma composição diferente. A par do azoto (N3) e do vapor de água (H2O), o metano (CH4) e o dióxido de carbono (CO2) eram abundantes. Menos abundantes seriam o monóxido de carbono (CO), o amoníaco (NH3) e o sulfureto de hidrogénio (H2S). O oxigénio, tão importante na atmosfera actual, estava quase completamente ausente.
As cinco etapas da biogénese
Foi neste contexto que se terá processado a biogénese, ou a formação da vida na Terra. É habitual considerar, neste processo, cinco etapas:
(1) a formação de uma atmosfera gasosa contendo as substâncias elementares para a formação da matéria orgânica (já referida) ;
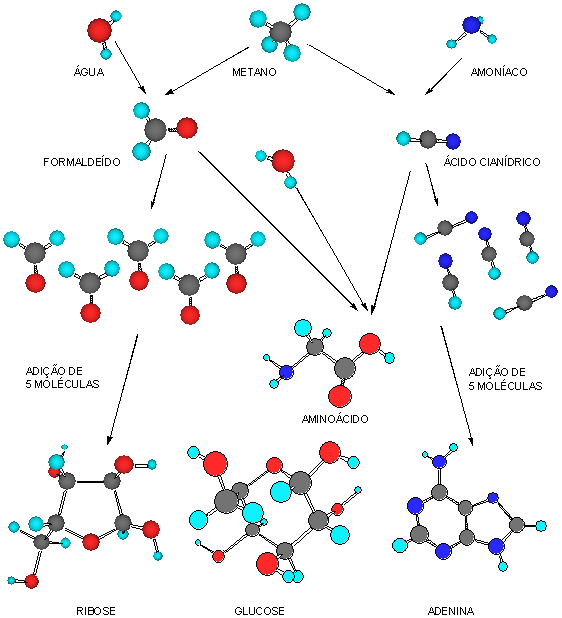
(2) a síntese dos monómeros biológicos, como os aminoácidos, os açúcares ou as bases azotadas;
(3) a polimerização destes monómeros e a formação das proteínas primitivas e das cadeias de ácidos nucleicos;
(4) a individualização de microgotas com uma identidade física e rudimentares faculdades metabólicas;
(5) o desenvolvimento de um mecanismo de reprodução que garantisse a transmissão da informação, aos descendentes.
a) Síntese de monómeros
A formação de aminoácidos e nucleótidos a partir de uma mistura de gases, redutora, com composição semelhante à da atmosfera primitiva, foi realizada experimentalmente, em 1952, por Harold Urey e Stanley Miller, recorrendo a descargas eléctricas como fonte de energia. Com esta espectacular experiência, Urey e Miller demonstraram ser possível a síntese de moléculas orgânicas por via não biótica e consolidaram a hipótese chave de Oparin e Haldane .
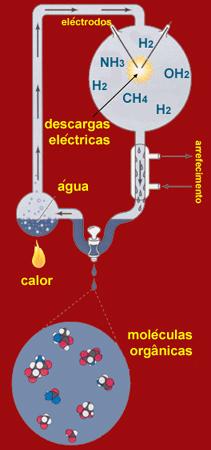
Aparato experimental de Miller
Desde logo, transpondo para as condições primitivas da Terra, passou a ser verosímil o cenário da síntese abiótica de pequenas moléculas orgânicas na atmosfera, sob a acção das fontes de energia disponíveis, nomeadamente as descargas eléctricas e as radiações ultravioletas mais energéticas que, devido à ausência total de camada de ozono, atingiam a superfície terrestre. Essas moléculas ter-se-iam acumulado em águas marinhas ou lacustres, ficando protegidas de reacções fotoquímicas posteriores e constituindo o caldo primordial, no entender de Haldane.
Sabe-se hoje que não terá sido essa a única fonte de moléculas orgânicas acumuladas. Também os cometas e os meteoritos que se esmagavam sobre a Terra transportariam consigo diversas moléculas orgânicas. Certos meteoritos, designados por condritos carbonados, contêm cerca de 3% do seu peso em carbono. Esse carbono encontra-se sob diversas formas, nomeadamente de aminoácidos, bases púricas e pirimídicas, e ácidos carboxílicos, o que prova que também fora da Terra seria possível a síntese de moléculas orgânicas.
b) Síntese de polímeros
A segunda etapa da biogénese trá sido a síntese dos polímeros que intervêm na organização da vida, por justaposição de monómeros sintetizados anteriormente. Grosso modo, podemos distinguir três tipos de polímeros:
- Uns, formam-se por ligação topo a topo, de uma série de aminoácidos; são as proteínas, que intervêm quer na estrutura dos organismos, quer no seu metabolismo como catalisadores.
- Outros, resultam da justaposição alternada de fosfatos e açúcares, aos quais se adicionam bases azotadas; são os ácidos nucleicos. Possuem faculdades catalíticas e são susceptíveis de autoduplicação.
- Outros ainda, formam estruturas ramificadas por associação de pequenos açúcares (monossacarídeos ou oligossacarídeos); são os polissacarídeos.

Síntese de polímeros
À luz da hipótese de Haldade, as reacções de polimerização teriam ocorrido, no “caldo primordial”, em consequência de uma grande acumulação de monómeros. Esta hipótese está hoje afastada, pelo motivo de que não se poderia explicar como é que uma reacção que liberta água (a polimerização) poderia ocorrer num ambiente com excesso de água. Sendo estas reacções reversíveis, um excesso de água deslocaria o equilíbrio no sentido da hidrólise, em lugar da polimerização. Para mais, as reacções de polimerização para se efectuarem necessitam de energia, o que não acontece com a hidrólise.
Existem duas maneiras de deslocar a reacção para a direita: (i) concentrar os reagentes, retirando a água produto da reacção; ou (ii) acoplando-a a outra reacção, “dadora” de energia.
Há boas razões para supor que alguns minerais tenham desempenhado um papel decisivo na “deslocação para a direita” destas reacções. Entre estes, merecem destaque os silicatos (argilas, micas, etc.), que são minerais com uma estrutura folheada e um elevado poder de adsorsão de moléculas. As argilas são referidas como “esponjas de moléculas”. No caulino, por exemplo, as camadas de silicatos distam entre si 0,71 nm, o que significa que um cubo de um centímetro de lado oferece uma superfície total de adsorsão de 2.800 m2( cerca de dois terços da superfície de um campo de futebol). Estes minerais, aliás, são conhecidos por apresentarem propriedades catalíticas e, como tal, são largamente empregues na indústria de síntese de compostos orgânicos.
Experimentalmente, Aharon Katchalsky demonstrou que um certo tipo de argilas promove a polimerização de cadeias polipeptídicas (análogas das proteínas) a partir de ésteres formados de aminoácidos e de adenosina monofosfato (adenilatos de aminoácidos) adsorvidos nas suas superfícies. Os adenilatos de aminoácidos são os precursores da síntese proteica em todos os organismos, pelo que é verosímil que mecanismos semelhantes tenham presidido à polimerização dos aminoácidos no período pré-biótico.
Poderá não ter sido, contudo, a única via possível para a síntese abiótica das proteínas. Sidney Fox mostrou que misturas secas de aminoácidos se polimerizam espontaneamente quando deixadas algumas horas a temperaturas da ordem de 130ºC. Chamou-lhes termoproteinóides. Em presença de polifosfatos e a temperaturas mais baixas, obtêm-se resultados comparáveis. Com base nos seus resultados experimentais, Fox formulou a hipótese de que os aminoácidos acumulados nos oceanos primitivos, porventura rejeitados sobre escórias vulcânicas, tenham sido polimerizados pelo calor. Retomadas posteriormente pelas águas, essas cadeias polipeptídicas poderão ter participado na organização dos primeiros protobiontes.
É importante chamar a atenção para o facto de que, qualquer que seja o processo abiótico de polimerização, ele conduz necessariamente a uma ordenação aleatória de aminoácidos, contrariamente ao que se verifica nos seres vivos.
Quanto aos ácidos nucleicos, a sua síntese abiótica, laboratorial, é tecnicamente acessível, em condições térmicas moderadas (55ºC), sendo possível conceber, para eles, um cenário semelhante ao da polimerização dos aminoácidos sobre superfícies de minerais adsorventes. Mas a ordenação dos nucleótidos será também aleatória.
c) A individualização de microgotas
Todos os organismos vivos são separados do meio em que vivem por uma barreira, que lhes confere a sua individualidade.
A formação de uma barreira individualizante terá sido obrigatoriamente um dos passos evolutivos em direcção às primeiras formas de vida. É verosímil que essas entidades primitivas tenham tido o aspecto de microgotas.
Entidades semelhantes foram criadas em laboratório por Oparin e por Fox, utilizando polímeros biológicos, proteínase e polissacarídeos, como a albumina e a goma arábica. Oparin mostrou que, espontaneamente, os polímeros em solução aquosa podem isolar-se do meio e formar coacervatos, isto é, microgotas ricas em polímeros, que ficam em suspensão na água. Por seu turno, Fox provocou a formação de um grande número de pequenas microsferas, dissolvendo na água proteinoides de origem abiótica. Nem os coacervatos de Oparin, nem as microsferas de Fox podem ser considerados como antepassados das células vivas. Os coacervatos e as microsferas constituem contudo um excelente material experimental, que Oparin e Fox exploraram para simulação de reacções metabólicas.
Uma das primeiras constatações de Oparin foi a de que os coacervatos eram entidades instáveis e de que a forma de evitar a sua perda era a de lhes proporcionar uma espécie de metabolismo primitivo.
Nas suas experiências, Oparin tirou partido do facto de que, num sistema com duas fases, as substâncias cuja solubilidade é diferente, consoante a fase, se concentram preferencialmente numa ou noutra. Juntando uma fosforilase (enzima da polimerização da glucose) a uma solução de histona (proteína) e goma arábica (polissacarídeo), Oparin não só deu lugar a que se formassem coacervatos, mas verificou ainda que o enzima se concentrou nestes últimos. Adicionando glucose-1-fosfato ao meio, constata-se que esta passa para o interior dos coacervatos e é polimerizada de seguida, pela fosforilase, em amido. A energia necessária à reacção provém da ligação fosfato da glucose-1-fosfato. O fosfato inorgânico libertado difunde para o exterior, como um dejecto. À medida que o fenómeno prossegue, os coacervatos aumentam de volume, devido à acumulação de amido.
A experiência pode ser repetida mas, desta vez, empregando dois enzimas: a fosforilase e uma amilase. Esta última catalisa a hidrólise do amido com libertação de maltose. Observa-se então que os coacervatos não aumentam de volume, porque o amido, à medida que é alongado pela fosforilase, é encurtado pela amilase.
Quando os coacervatos se tornam grandes de mais, dividem-se espontaneamente em coacervatos "filhos". Entre estes, aqueles que tiverem ficado com moléculas de fosforilase continuarão a poder polimerizar a glucose, a crescer e a dividir-se. Os outros não terão possibilidade de subsistir. Para que o fenómeno se perpetuasse como numa célula, só faltaria um sistema que sintetizasse a fosforilase, com aquela composição exacta e nas quantidades necessárias !
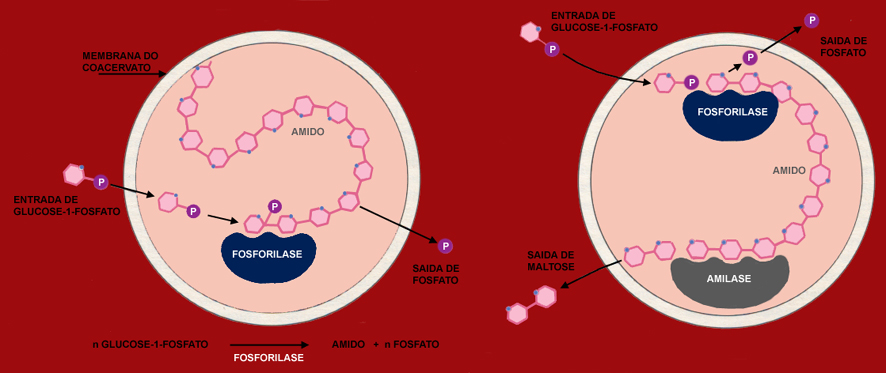
Experiência de Oparin
Nos coacervatos de Oparin, são os polissacarídeos que formam a barreira limitante e os agentes catalisadores são incorporados a posteriori. Nas microsferas de Fox, são os próprios proteinóides que asseguram a constituição da “membrana” e actuam como enzimas das reacções, podendo catalisar diversas reacções, nomeadamente a decomposição da glucose.
As experiências de Oparin e de Fox mostraram que :
(1) a formação de emulsões de microgotas (coacervatos ou microsferas) corresponde a uma tendência natural dos polímeros em solução;
(2) os eventos que neles decorrem, fundamentam-se nas leis da química física;
(3) a evolução de entidades tão simples, mas com características novas, como os protobiontes, pode ter-se baseado unicamente nas suas características físico-químicas, seleccionadas exclusivamente em função da sua capacidade de sobrevivência.
d) Um mecanismo de replicação
Os protobiontes atrás referidos estão ainda muito longe dos seres vivos mais simples. Falta-lhes a capacidade de reprodução, isto é, o “plano de construção” que possa ser copiado e utilizado para fabricar outro microssistema idêntico ao primeiro. Este é na realidade o fosso que separa o protobionte, capaz de alguns actos metabólicos, de crescer e de se dividir, do eubionte, verdadeiro ser vivo, capaz de se reproduzir, gerando seres idênticos a si próprio, com as mesmas faculdades bioquímicas.
A grande diferença reside em ter ou não ter informação.
A grande dificuldade reside em compreender como se estabeleceu a correspondência proteína/ácido nucleico. Sabemos que em todos os organismos vivos actuais a sequência de nucleótidos de alguns ácidos nucleicos determina a sequência dos aminoácidos na proteína, isto é, a sua estrutura primária. Mas também são proteínas (com uma estrutura primária definida) que são responsáveis pela catálise da síntese dos ácidos nucleicos como ainda, do estabelecimento das ligações peptídicas entre os diferentes aminoácidos.
Existem bons argumentos que sustentam a proposta de Gilberto, segundo a qual o RNA teria podido desempenhar a dupla função, de codificar a síntese de proteínas e de actuar cataliticamente ao nível da sua própria síntese. Dito de outra forma, a vida primitiva ter-se-ia baseado exclusivamente no RNA, sem o DNA nem enzimas interventores na replicação dos ácidos nucleicos e na síntese proteica. O sistema que conhecemos actualmente, baseado no DNA e em enzimas proteicos, e que é comum a todos os seres vivos, teria surgido posteriormente, seleccionado pelas vantagens que oferece para a propagação da vida.
________________________________________________________________________________________________________________________