ENERGIA
I - METABOLISMO E ENERGIA
Os organismos vivos, quer sejam monocelulares ou pluricelulares, são capazes de desenvolver actividades diversas, como crescimento, movimento, secreção de substâncias, produção de electricidade, de luz, etc. Essas actividades são redutíveis a reacções químicas que, na sua globalidade, recebem a designação genérica de metabolismo. Considera-se, por sua vez, que o metabolismo de uma célula ou de um organismo é formado essencialmente por dois grupos de reacções químicas:
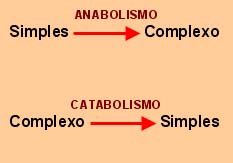
Anabolismo, que agrupa as reacções químicas cuja função consiste na elaboração de moléculas complexas a partir de elementos ou moléculas simples; o catabolismo, que agrupa um conjunto de reacções conducentes à degradação química de moléculas complexas, em produtos mais simples.
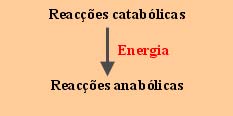
Numa célula, as reacções anabólicas e catabólicas decorrem geralmente em simultâneo. Este facto levanta alguns problemas, quando se encaram as reacções numa perspectiva energética. Com efeito, as reacções anabólicas são consumidoras de energia, enquanto que as reacções catabólicas, produzem-na. O problema consiste em saber como é que uma célula capta energia de uma reacção catabólica para a utilizar numa reacção anabólica.
II - ENERGIA LIVRE E ENTROPIA: A SEGUNDA LEI DA TERMODINÂMIVA
Antes de avançarmos, é necessário conhecer as condições que determinam se uma reacção química pode ou não, ocorrer. Para tal, recorramos a uma analogia mecânica.
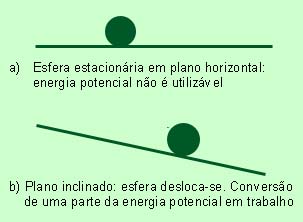
Considere-se um corpo perfeitamente liso, uma esfera, por exemplo, colocada num plano horizontal, igualmente liso. A esfera possui uma certa energia potencial; todavia inutilizável enquanto o plano se mantiver horizontal. Inclinemos o plano. A esfera desliza . Ao deslocar-se, a esfera executa um trabalho; a quantidade de trabalho produzido é igual à quantidade e energia potencial perdida.
Teoricamente, seria possível imaginar um corpo deslizando num plano de comprimento infinito, até que toda a energia potencial fosse convertida em trabalho. Na realidade, contudo, assim não se passa: só uma parte da energia potencial é efectivamente utilizável para produzir um trabalho. Esta fracção de energia, denomina-se energia livre e representa-se por G. Consequentemente:
a energia livre
é a fracção da energia potencial convertível em trabalho
Sabemos, por experiência, que um corpo nunca sobe espontaneamente um plano inclinado. Este movimento só é possível mediante um investimento em energia. Abandonado a si próprio, o corpo deslizará no plano e a energia livre decrescerá à medida que se converte em trabalho.
Para melhor precisarmos estas noções, pensemos no caso de uma máquina térmica, à qual se fornece uma certa quantidade de energia Q, à temperatura T1, sendo conduzida, em seguida a uma temperatura T2.
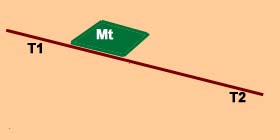
A quantidade de trabalho W fornecida por esta máquina é, como se sabe, dado pela fórmula:
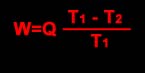
de onde se deduz que
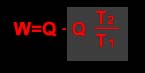
Desta última fórmula se deduz que o máximo trabalho realizado será sempre inferior ao trabalho total possível de uma quantidade Q T2/T1, ou, o que o mesmo, T2Q/T1. Uma tal máquina não poderá transformar totalmente a energia em trabalho, sem que T2 seja igual a zero (zero absoluto). Para todo T2>0 , uma parte da energia total não será portanto disponível.
Esta quantidade de energia não disponível a uma temperatura T é determinada pelo produto T Q/T1, em que Q/T1 se representa pela letra S e de denomina entropia.
Portanto,
a entropia
mede a fracção de energia total que é inutilizável para efectuar um trabalho
A segunda lei da termodinâmica exprime a tendência para a desordem que se verifica em qualquer sistema composto por um certo número de partículas. Como aliás a experiência nos ensina, as coisas deixadas ao abandono não se organizam espontaneamente: tal como as moléculas de açúcar no fundo de uma chávena de café tenderão a espalhar-se (mesmo sem que se agite com a colher), também os papeis na secretária não se arrumarão espontaneamente (antes pelo contrário), nem as peças de um carro se rejuvenescerão sem que se invista trabalho. Portanto, um sistema abandonado a si mesmo tende a passar de um estado organizado, para um estado desorganizado. Contrariar esta tendência obriga a um investimento de trabalho.
Quanto maior for a fracção de energia não mobilizável para trabalho, maior será a desordem. Assim se compreende que a entropia mede também o estado de desordem. Quanto maior for a desordem, maior será a entropia. A entropia máxima reflecte portanto o estado de desordem máximo.
Todos os fenómenos naturais mostram um aumento inexorável de entropia. A vida distingue-se pelo facto de o estado de entropia máximo, isto é, a morte, ser retardado, mas não abolido, por um consumo de energia a partir do meio.
Aplicando o raciocínio a qualquer outro sistema que não a uma máquina térmica, poderemos descrever mais genericamente um sistema funcionando a uma temperatura T :
em que G é a energia livre, H, a entalpia ( ou energia calórica total ) e S, a entropia.
Com excepção de T, é impossível determinar os valores absolutos destas variáveis, para o que seria necessário conhecer a energia livre e a entropia do universo. Podemos todavia determinar as variações sofridas por um sistema quando se passa de um estado inicial a um novo estado, e escrever
DG = DH - TDS
Num sistema bioquímico, como em qualquer outro, qualquer mudança ocorre sempre no sentido de uma diminuição da energia livre
Por exemplo, uma reacção só pode realizar-se espontaneamente se o resultado se traduzir numa diminuição de energia livre. Por sua vez, uma reacção que implique um aumento de energia livre, não poderá ocorrer se não dispuser de uma fonte adequada de energia.
As reacções químicas são acompanhadas quer por uma produção de calor (exotérmicas), quer por uma absorção de calor (endotérmicas). A variação térmica não corresponde, contudo, a uma modificação da energia livre, pois para isso seria necessário conhecer a variação de entropia. É portanto a variação da energia livre, e só essa, que nos permitirá dizer se uma reacção química é possível ou não.
Uma reacção química é designada endergónica se a variação de energia livre for positiva; significa então que será necessário fornecer energia para que ela se realize. Pelo contrário, são exergónicas as reacções às quais corresponde uma variação negativa da energia livre. Isto é, no decurso da reacção, uma certa quantidade de energia livre, é libertada.
Uma forma de fornecer energia a uma reacção endergónica consiste em a acoplar a uma reacção exergónica. A única imposição termodinâmica para que uma reacção exergónica possa fazer funcionar uma reacção endergónica é que a reacção exergónica forneça mais energia livre do que a necessária à reacção endergónica. É necessário pois que, globalmente, a reacção total seja exergónica
Numa reacção química do tipo
![]()
o que se pode medir directamente é DH, a variação de energia calórica total. Quanto a DG, demonstra-se que:
DG = DGº + R T ln [C] [D] / [A] [B]
onde DGº é a energia livre padrão, dada pela fórmula
DGº = - R T ln K
sendo K a constante de equilíbrio da reacção, R, a constante dos gases perfeitos e T, a temperatura absoluta.
Vimos acima que uma reacção química será termodinamicamente possível, se for exergónica. O facto de ocorrer ou não, é outra questão, pois, tal como um corpo não deslizará num plano inclinado rugoso, onde o atrito é grande, também uma reacção química poderá não se iniciar espontaneamente, apesar de estarem reunidas as condições termodinâmicas.
Por exemplo, a sacarose em solução em meio neutro, não sofrerá uma reacção espontânea de hidrólise, ainda que esta seja termodinamicamente possível, porque as moléculas não se encontram em estado reactivo. Alternativamente, se se adicionar um enzima específico, o açúcar será hidrolisado (ver enzimas) .
III - REACÇÕES DE OXI-REDUÇÃO
De tudo quanto se disse, emerge o problema de saber de que modo a célula capta energia de uma reacção catabólica para a utilizar numa reacção anabólica. Contudo, o problema pode ser equacionado através de três questões:
a) qual é a fonte biológica de energia ?
b) de que modo a energia é captada pela célula ?
c) de que modo é a energia utilizada para efectuar um trabalho ?
Numa célula, a energia necessária às reacções endergónicas, entre as quais se situam as que visam a transformação dos alimentos em materiais celulares, é fornecida, essencialmente, por reacções de oxi-redução (redox).
Se bem que originalmente a oxidação tenha sido considerada como um fenómeno resultante da combinação de um elemento ou de uma molécula com o oxigénio, o conceito de oxidação é hoje definido de modo mais amplo e engloba os fenómenos de perda de electrões. O fenómeno inverso, no qual um elemento ou uma molécula ganha electrões, é designado por redução.
Assim sendo, toda a reacção de oxidação encontra-se obrigatoriamente acoplada a uma reacção de redução. Por conseguinte, qualquer reacção de transferência de electrões, implica a participação de um oxidante (agente captador de electrões) e de um redutor (agente dador de electrões) Por esta razão, se refere correntemente a reacções de oxi-redução ou redox.
Por exemplo, na situação seguinte, Fe+++ actua como oxidante que, ao captar um electrão, é reduzido em Fe++ ; pelo contrário, Fe++ actua como redutor que, ao libertar um electrão, é oxidado em Fe+++.
Fe++ <===> Fe+++ + e-
Portanto:
aquele que se oxida é o redutor; aquele que se reduz, é o oxidante
Os átomos e as moléculas diferem quanto à afinidade para os electrões. Assim, por exemplo, o bromo é um oxidante suficientemente poderoso para libertar o iodo do ião iodeto, “roubando” a este 2 electrões:
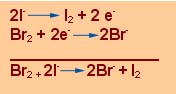
Pela mesma razão, uma lâmina de zinco mergulhada numa solução de um sal de cobre, recobre-se de uma camada de cobre: o zinco captura electrões do ião cobre e precipita o cobre metálico, enquanto que o zinco reduzido em ião Zn++ se dissolve. Pelo contrário, se se mergulhar uma lâmina de cobre numa solução de um sal de zinco, nada acontece.
A transferência de electrões entre substâncias com diferentes afinidades para estas partículas, está na origem das pilhas eléctricas. Com efeito, se, numa solução contendo simultaneamente iões Zn e Cu, mergulharmos duas barras, uma de zinco e outra de cobre, verificar-se-a o desenvolvimento de uma diferença de potencial eléctrico entre as duas barras metálicas (eléctrodos). A pilha assim constituída desenvolve uma força electromotriz de 1,1 volts, aproximadamente.
A capacidade de um átomo ou uma molécula "roubarem" electrões a outros,oxidando-os, designa-se por potencial de oxido-redução ou potencial redox.
Entre as diversas substâncias com aptidão para “roubar” electrões a outras, o oxigénio ocupa uma posição de destaque. Daí a designação de oxidação dada genericamente a estes fenómenos.
No caso das moléculas orgânicas, o fenómeno de oxidação é geralmente sinónimo de desidrogenação. Isto é, por cada electrão perdido, a molécula perde também um protão. Neste casos, a situação "complica-se" pois não só será necessário acoplar a reacção a um redutor (receptor de electrões), mas também a um outro agente, captador de protões. Tratando-se de reacções de desidrogenação, serão catalisadas por desidrogenases.
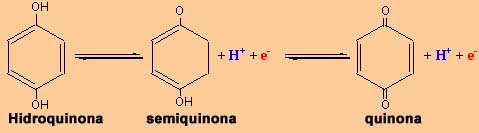
Oxidação de uma molécula orgânica, a hidroquinona
Como etapa intermédia, forma-se uma semiquinona
As desidrogenases são enzimas que catalisam a desidrogenação de substratos. São específicas dos substratos e encontram-se sempre associadas a coenzimas, transportadores de protões. Entre os mais comuns, referem-se o NAD (Nicotinamida Adenina Dinucleótido), o NADP, o FAD (Flavina Adenina Dinucleótido), a FMN (Flavina Mononucleótido).
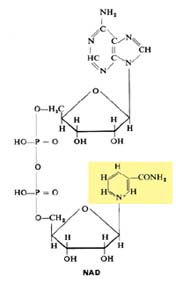
NAD
No caso do NAD, a componente interveniente é a nicotinamida, assinalada em fundo amarelo na imagem precedente, onde o N se encontra, na realidade, sob forma catiónica (+). O NAD+ é reduzido fixando um electrão e um átomo de hidrogénio:
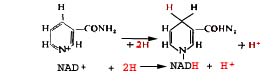
Por simplificação, escreve-se habitualmente:
NAD + 2H ---> NADH2
O FAD, outro dos coenzimas das desidrogenases, pelo contrário, é capaz de fixar simultaneamente dois electrões e dois
protões: FAD + 2H ---> FADH2
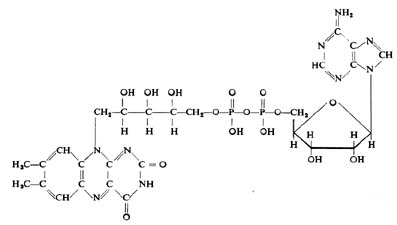
FAD
Em conclusão, a maioria das reacções de oxidação que ocorrem na célula, implicam a existência, para além do substrato que será oxidado, de uma desidrogenase específica dessa oxidação e um aceitador de protões, que será necessariamente reduzido.
IV - O PAPEL DOS FOSFATOS
Como se disse, a energia necessária para a actividade anabólica provem das reacções catabólicas. Este facto conduz à noção de acoplamento, que permite explicar o modo pelo qual a energia catabólica é utilizada nas sínteses e nas restantes actividades celulares.
Se bem que as reacções exergónicas libertem sempre uma fracção de energia sob a forma de calor, não será este utilizado no trabalho celular. O que interessa nesta perspectiva, é a energia livre utilizável para sintetizar moléculas. Contudo, qualquer que seja a natureza dos materiais oxidados (lípidos, glúcidos, aminoácidos, etc.) a energia livre tornada disponível pelas oxidações, não é directamente empregue nos processos endergónicos; ela será empregue na síntese de uma classe especial de compostos intermédios que constituem o “combustível” correntemente utilizado nos processos anabólicos (endergónicos).
Esta síntese efectua-se por incorporação de um radical R a um dos reagentes. No decurso da evolução, operou-se uma selecção que incidiu sobre a selecção desse radical como também sobre a natureza das moléculas capazes de realizar essas reacções.
O radical R mais geralmente utilizado é o resíduo fosforilo, o qual, nas condições de pH próprias do meio celular, se encontra sob a forma aniónica (forma B).

Resíduos fosforilo
O dador principal deste radical é a adenosina trifosfato ou ATP. Não sendo o único, o ATP é todavia o membro mais notório de uma família de compostos fosforilados dotados de um elevado potencial energético. Ou, dito de outro modo,
são compostos cuja hidrólise disponibiliza uma energia livre excepcionalmente grande.
Com efeito, enquanto que a hidrólise dos fosfatos comuns liberta cerca de 3 Kcal M-1, a dos os fostatos “ricos em energia”, como o ATP, aproxima-se de 8 Kcal M-1.
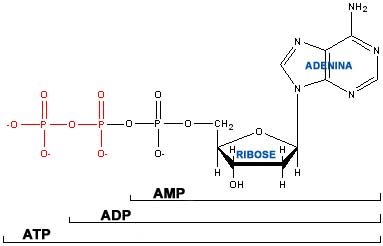
Adenosina trifosfato (ATP)
No caso do ATP, a hidrólise liberta um fosfato inorgânico (Pi) e dá origem à adenosina difosfato ou ADP. Esta, por sua vez, pode ser hidrolisada em adenosina monofosfato ou AMP. Ambas as hidrólises são acompanhadas pela libertação de 8 Kcal M-1. Pelo contrário, a hidrólise do AMP não liberta senão 3 Kcal M-1.

V- ACOPLAMENTOENERGÉTICO
Em virtude da primeira lei da termodinâmica, a energia das reacções catabólicas deve ser utilizada na actividade anabólica da célula. O fundamento da energética bioquímica repousa no conceito de acoplamento energético. Como se viu, a única exigência do ponto de vista termodinâmico, para que uma reacção exergónica possa ser utilizada na viabilização de uma reacção endergónica, é que liberte mais energia do que aquela que será consumida pela segunda.
Do ponto de vista químico, a realização de acoplamento implica que a transferência de energia de uma reacção à outra se faça por intermédio de um reagente comum às duas reacções. Os compostos “ricos em energia” atrás referidos, designadamente o ATP, constituem, de facto, os reagentes comumente empregues pelo metabolismo celular. O exemplo seguinte, relativo à síntese do glicogénio, ilustra este fenómeno de acoplamento energético.
O glicogénio é um composto polimérico de moléculas de glucose ligadas entre si por pontes glicosídicas (ver glúcidos). Antes de se adicionar às outras, cada molécula de glucose deve ser objecto de uma fosforilação. Todavia, esta operação é endergónica, pelo que deverá ser conjugada a uma reacção exergónica:

O balanço salda-se por uma variação de energia livre de - 5 Kcal M-1, o que indica que a reacção se fará espontaneamente, em presença do enzima específico, uma hexoquinase.
____________________________________________________________________________________________________
RETORNO >